

-
摘要:
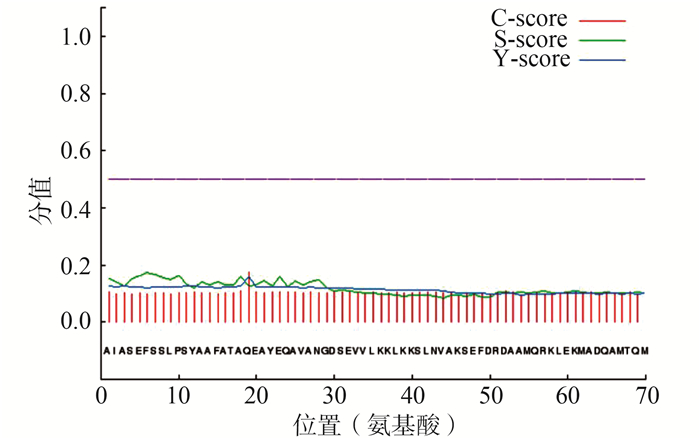
目的 SARS-CoV-2感染不仅引起呼吸系统病变,且可能累及生殖系统及中枢神经系统。本研究探讨SARS-CoV-2 nsp8基因变异、Nsp8蛋白结构与生物学功能及靶向药物,为建立更有效的防控策略奠定基础。 方法 采用生物信息学技术和大型生物数据库,分析nsp8基因变异性、Nsp8理化特征、空间结构、抗原表位、生物功能、药物结合靶点等。 结果 基于3种28株冠状病毒的nsp8序列构建了进化树;SARS-CoV-2毒株间nsp8基因保守性为99%~100%,与SARS遗传距离较MERS近;Nsp8无信号肽及跨膜区;在体外网织红细胞内半衰期为4 h,具有亲水性;获得Nsp8二级和三级结构模型;Nsp8具有线性B细胞和CTL细胞抗原表位、磷酸化及SUMO修饰位点;从DrugBank数据库筛选出可靶向结合Nsp8的四种药物。 结论 Nsp8具有典型抗原分子特征,参与病毒复制,且同种不同株nsp8基因高度保守,在病毒致病机制、分型及防治研究中具有应用潜力。在全球亟需新型冠状病毒肺炎特效防治措施的状况下,靶向结合Nsp8的化疗药物具有重要科学意义和应用价值。 Abstract:Objective To explore SARS-CoV-2 nsp8 genetic variation, Nsp8 protein structure, biological function and targeted drugs, and to lay foundation for establishing more effective prevention and control strategies. Methods Analyses of nsp8 genetic variability, physical and chemical characteristics, spatial structure, antigenic epitopes, biological function, and drug combined targets of Nsp8 were carried out using bioinformatics technology and large biological databases. Results Based on nsp8 sequences of 28 isolates of coronavirus of three species, evolutionary tree was successfully constructed. SARS-CoV-2 isolates showed 99%-100% conservation of nsp8 genes, less genetic distance to SARS than MERS isolates. Nsp8 had no signal peptide and transmembrane area. In reticulocytes in vitro, Nsp8 had a half-life of 4 h and was hydrophilic. A secondary model and a tertiary structure model were established. Linear B cell and CTL antigenic epitopes, phosphorylation and SUMB modification sites were found in Nsp8. Using the DrugBank database, four drugs targeted Nsp8 were obtained. Conclusions Nsp8 possesses the characteristics of typical antigens, participates in viral replication, and various isolates of the same species share high conservation of nsp8 gene, suggesting potential applications in researches on pathogenic mechanism, genotyping and prevention of this virus. Notably, this is the first report on Nsp8-targeted chemotherapeutic drugs, and the findings can be of considerable scientific significance and application value, under the conditions that measures with special effect for COVID-19 prevention and control are urgently needed. -

图 1 SARS-CoV-2、SARS及MERS病毒株nsp8基因遗传进化树
Figure 1. Genetic evolutionary tree of nsp8 genes of SARS-CoV-2, SARS and MERS isolates

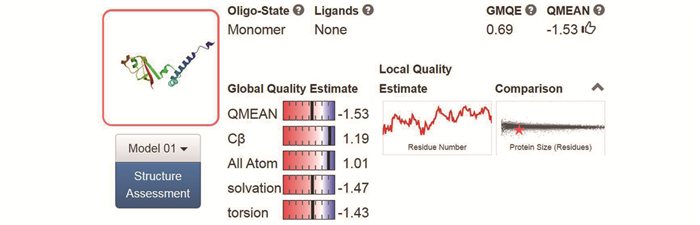
图 9 Nsp8三级结构的SWISS-MODEL分析
Figure 9. Analysis of tertiary structure of Nsp8 using SWISS-MODEL

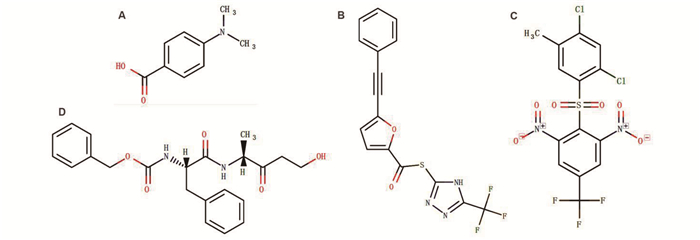
图 11 基于DrugBank的Nsp8靶向结合药物分子分析
注:A:4-(Dimethylamino) benzoic acid, C9H11NO2, DB08748;B:S-[5-(TRIFLUOROMETHYL)-4H-1, 2, 4-TRIAZOL-3-YL] 5-(PHENYLETHYNYL) FURAN-2-CARBOTHIOATE, C16H8F3N3O2S, DB07743;C:2-[(2, 4-DICHLORO-5-METHYLPHENYL)SULFONYL]-1, 3 -DINITRO-5- (TRIFLUOROMETHYL) BENZENE, C14H7Cl2F3N2O6S, DB07620; D:NALPHA-[(BENZYLOXY)CARBONYL]-N-[(1R)-4-HYDROXY-1-METHYL-2-OXOBUTYL] -L-PHENYLALANINAMIDE, C22H26N2O5, DB08732。
Figure 11. Analysis of Nsp8-targeted drugs based on DrugBank
表 1 冠状病毒SARS-COV-2、SARS和MERS毒株Nsp8比较
Table 1. Comparison of Nsp8 proteins of coronavirus SARS-COV-2, SARS and MERS strains
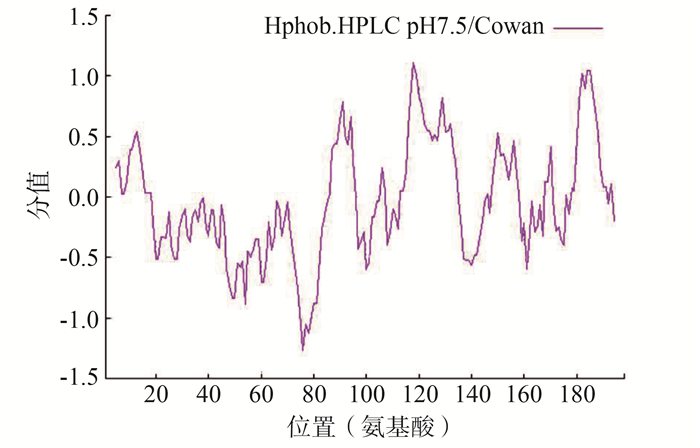
属性 病毒株 SARS-COV-2/CHN/ Wuhan-Hu-1/2020 SARS/CHN/GD01/2003 MERS/KOR/066-2015/2015 基因结构 nsp8基因为多聚蛋白基因ORF1ab中一段编码区,长594 bp,GC含量38.38% nsp8基因为多聚蛋白基因ORF1ab中一段编码区,长594bp,GC含量42.08% nsp8基因为多聚蛋白基因ORF1ab中一段编码区,长597 bp,GC含量41.71% 分子质量(Dal) 21 881.08 21 866.03 21 886.17 氨基酸数量(aa) 198 198 199 等电点 6.58 6.58 6.83 半衰期(h) 4.4 4.4 4.4 不稳定系数 37.78 39.23 25.77 脂肪族指数 88.33 87.83 95.63 平均亲水系数 -0.192 -0.226 -0.101 Nsp8二级结构 α螺旋54.04%、随机卷曲22.22%、β转角6.57%和延伸链17.17% α螺旋52.02%、随机卷曲21.21%、β转角6.06%和延伸链20.71% α螺旋53.27%、随机卷曲23.12%、β转角6.03%和延伸链17.59% Nsp8三级结构 


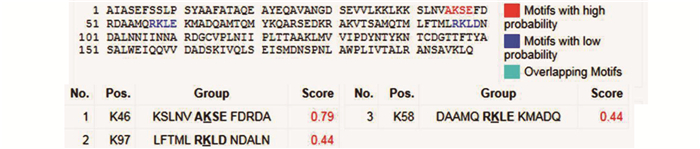
磷酸化位点 15个丝氨酸、13个苏氨酸和6个酪氨酸磷酸化位点 14个丝氨酸、12个苏氨酸和7个酪氨酸磷酸化位点 15个丝氨酸、11个苏氨酸和5个酪氨酸磷酸化位点 糖基化位点 无O-糖基化位点 3个O-糖基化位点 无O-糖基化位点 B细胞抗原表位(长度≥9 aa) 4个B细胞表位:23~33、50~58、73~81、158~179 aa 4个B细胞表位:23~33、50~58、73~81、158~178 aa 4个B细胞表位:23~34、50~59、74~82、159~179 aa HLA-A*02∶01限制性CTL细胞表位(长度=9 aa,分值≥20) 14个CTL表位:90~98、152~160、1~9、25~33、34~42、102~110、151~159、180~188、54~62、123~131、179~187、86~94、118~126、161~169 aa。 与SARS-COV-2/CHN/ Wuhan-Hu-1毒株相同 6个CTL表位:91~99、103~111、115~123、35~43、38~46、159~167 aa Nsp8靶向结合药物 DrugBank Accession No. DB08748、DB07620、DB07743、DB08732 与SARS-COV-2/CHN/ Wuhan-Hu-1毒株相同  下载: 导出CSV
下载: 导出CSV
-
[1] Li Y, Zhou W, Yang L, et al. Physiological and pathological regulation of ACE2, the SARS-CoV-2 receptor[J]. Pharmacol Res, 2020, 157: 104833. DOI: 10.1016/j.phrs.2020.104833. [2] Yadav PD, Potdar VA, Choudhary ML, et al. Full-genome sequences of the first two SARS-CoV-2 viruses from India[J]. Indian J Med Res, 2020, 151(2&3): 200-209. DOI: 10.4103/ijmr.IJMR_663_20. [3] Li MY, Li L, Zhang Y, et al. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues[J]. Infect Dis Poverty, 2020, 9(1): 45. DOI: 10.1186/s40249-020-00662-x. [4] Zheng J. SARS-CoV-2: an emerging coronavirus that causes a global threat[J]. Int J Biol Sci, 2020, 16(10): 1678-1685. DOI: 10.7150/ijbs.45053. [5] Payus AO, Liew Sat Lin C, Mohd Noh M, et al. SARS-CoV-2 infection of the nervous system: a review of the literature on neurological involvement in novel coronavirus disease-(COVID-19)[J]. Bosn J Basic Med Sci, 2020, 20(3): 283-292. DOI: 10.17305/bjbms.2020.4860. [6] Ahsan W, Javed S, Bratty MA, et al. Treatment of SARS-CoV-2: How far have we reached?[J]. Drug Discov Ther, 2020, 14(2): 67-72. DOI: 10.5582/ddt.2020.03008. [7] Li H, Zhou Y, Zhang M, et al. Updated approaches against SARS-CoV-2[J]. Antimicrob Agents Chemother, 2020, 64(6): e00483-e00520. DOI: 10.1128/AAC.00483-20. [8] Yang D, Leibowitz JL. The structure and functions of coronavirus genomic 3' and 5' ends[J]. Virus Res, 2015, 206: 120-133. DOI: 10.1016/j.virusres.2015.02.025. [9] Gasteiger E, Hoogland C, Gattiker A, et al. Protein identification and analysis tools on the ExPASy server[M]. Totowa: Humana Press, 2005: 571-607. [10] Petersen TN, Brunak S, Heijne GV, et al. SignalP 4.0: discriminating signal peptides from transmembrane regions[J]. Nature Methods, 2011, 8(10): 785-786. DOI: 10.1038/nmeth.1701. [11] Blom N, Sicheritz-Ponten T, Gupta R, et al. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence[J]. Proteomics, 2004, 4(6): 1633-1649. DOI: 10.1002/pmic.200300771. [12] Steentoft C, Vakhrushev SY, Joshi HJ, et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology[J]. EMBO J, 2013, 32(10): 1478-1488. DOI: 10.1038/emboj.2013.79. [13] Li YC, Bai WZ, Hashikawa T. The neuroinvasive potential of SARS-CoV2 may play a role in the respiratory failure of COVID-19 patients[J]. J Med Virol, 2020, 92(6): 552-555. DOI: 10.1002/jmv.25728. [14] Holtmann N, Edimiris P, Andree M, et al. Assessment of SARS-CoV-2 in human semen-a cohort study[J]. Fertil Steril, 2020, 114(2): 233-238. DOI: 10.1016/j.fertn. [15] Mary B, Maurya S, Arumugam S, et al. Post-translational modifications in capsid proteins of recombinant adeno-associated virus (AAV) 1-rh10 serotypes[J]. FEBS J, 2019, 286(24): 4964-4981. DOI: 10.1111/febs.15013. [16] Vargab B, Sujoy B. Immunoinformatics-aided identification of T cell and B cell epitopes in the surface glycoprotein of 2019-nCoV[J]. J Med Virol, 2020, 92(5): 495-500. DOI: 10.1002/jmv.25698. [17] Aminikhah M, Yekaninejad MS, Nicknam MH, et al. HLA Class Ⅰ and Class Ⅱ genes distribution of the Iranian Sistanis[J]. Iran J Immunol, 2018, 15(2): 97-111. http://www.ncbi.nlm.nih.gov/pubmed/29947339 [18] Saxena A. Drug targets for COVID-19 therapeutics: ongoing global efforts[J]. J Biosci, 2020, 45(1): 87. DOI: 10.1007/s12038-020-00067-w. -



 点击查看大图
点击查看大图

计量
- 文章访问数: 322
- HTML全文浏览量: 338
- PDF下载量: 42
- 被引次数: 0